LysoSensor®系列探针是主要对活细胞中的酸性区室(如溶酶体、顶体精子)动态合成和功能进行研究的一类荧光探针,该类探针的染色具有向酸性,可通过质子化作用在酸性细胞器内累积。该类探针的这种质子化作用还可解除染料的荧光淬灭性,使其所发射的荧光强度更强。因此,与LysoTracker系列探针相比,LysoSensor®系列探针随着细胞器的酸化程度其荧光强度呈pH依赖型(向酸)增强。
本品LysoSensor™ Green DND-189具有较低的pKa值,仅在酸性区室内才具有荧光。本品以溶于无水DMSO的1 mM储存液形式提供。
如对活细胞中的酸性区室进行选择性标记和染色,可选择LysoTracker®系列探针。
| 分子式(Molecular Formula) | C24H22N4O2 |
| 分子量(Molecular Weight) | 398.46 |
| Ex/Em | 443/505 nm |
| pKa | ~5.2 |
| 颜色(Color) | 绿色 |
| 结构式(Structure) | 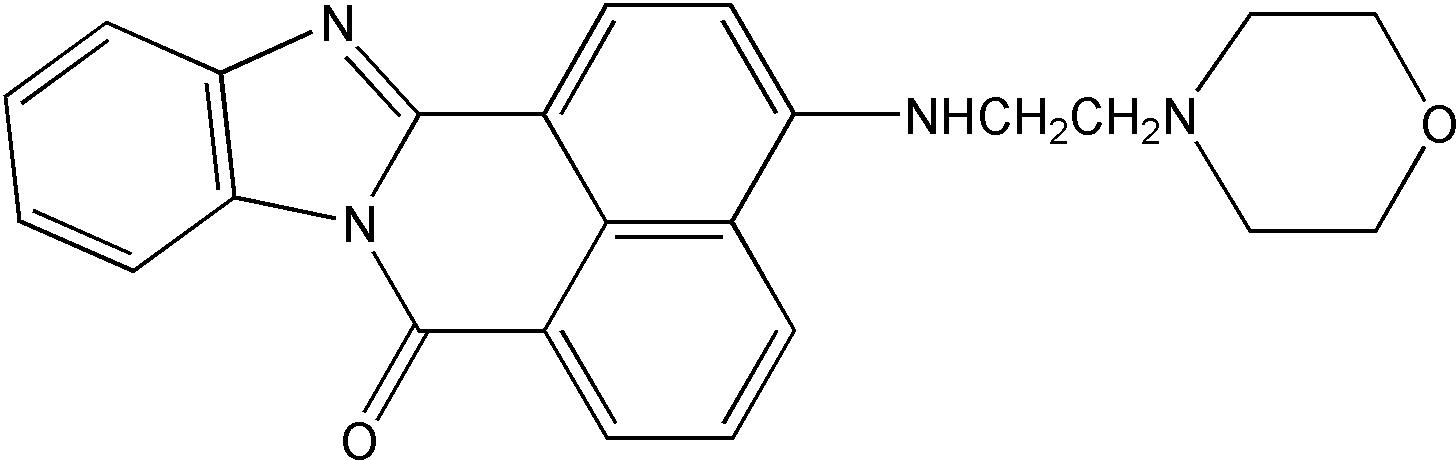 |
冰袋运输。-20℃避光干燥保存,避免反复冻融,有效期6个月。
Q:保质期多久?
A:-20℃避光保存,半年有效。
Q:四种溶酶体探针的区别?
A:LysoTracker®--不依赖 pH 的活细胞溶酶体荧光探针(40738 和 40739); LysoSensor™--依赖 pH 的活细胞溶酶体荧光探针(40767 和 40768)。
Q:染色时细胞的状态?
A:这四种探针是对活细胞中的酸性区室进行染色,所以必须要在活细胞状态下染色。
Q:染色完成后,细胞是否能固定后再检测荧光信号?
A:固定后会破坏溶酶体的酸性环境从而会造成荧光减弱甚至消失的情况,染色前后均不建议固定。
Q:可以和二抗一起孵育吗?
A:不可以,孵育二抗必须要透化处理,而透化会破坏溶酶体的酸性环境,影响染色效果。
Q:这个探针可以染自噬溶酶体吗?可以染色其他酸性细胞器吗?
A:自噬溶酶体和正常的溶酶体在形态结构上有较大差异,该探针主要是针对常规的溶酶体染色,对于自噬溶酶体的染色效果不是很确定。可以参考一些使用过该产品发表的文献资料确定可以染色其他酸性细胞器。
Q:植物细胞和组织适用吗?
A:理论上可以,一般植物细胞或者组织要制成原生质体。
Q:染色后可以放置一夜,再荧光检测吗?
A:建议染色完立即检测荧光,时间过长,荧光信号会逐渐减弱。
Q:用细胞培养基配成工作液以后稳定吗?可以存放多久?
A:工作液是需要现配现用的,不建议保存。
Q:可以使用的检测仪器有哪几种?
A:荧光显微镜、共聚焦显微镜、荧光酶标仪、流式细胞仪。
Q:如何做到同时染色溶酶体和细胞核的?
A:建议用 Hoechst 33258/ Hoechst 33342 染色细胞核,这个染料可以直接染活细胞不需要固定操作。
Q:荧光信号较弱?
A:大致有 3 个因: 1.溶酶体探针浓度过低,建议适当增大浓度; 2.染色时间过短,建议延长染色时间; 3.染色完之后,放置过长时间才检测信号;建议染色完立刻检测。
Q:有哪些原因会导致没有荧光信号?
A:大致有 3 个原因: 1.先用多聚甲醛、乙醛等固定细胞,破坏了溶酶体的酸性环境。 2.染色完后,又用多聚甲醛固定细胞,或者透化细胞,破坏了溶酶体的酸性环境;染色前后均不建议固定细胞; 3.染色完到上机检测这一段时间过长,并且没有避光处理。建议避光染色,染色完立即检测荧光信号。
Q:在活细胞状态下,用 50 µM 探针染色 50 min,然后发现除了溶酶体,细胞质中也有荧光信号,这主要是什么原因导致?
A:1.溶酶体探针浓度过大,建议适当降低浓度;
2.溶酶体探针染色时间过长,建议适当缩短时间。
[1] Yan JM, Zhang WK, Yan LN, Jiao YJ, Zhou CM, Yu XJ. Bunyavirus SFTSV exploits autophagic flux for viral assembly and egress. Autophagy. 2022;18(7):1599-1612. doi:10.1080/15548627.2021.1994296(IF:16.016)
[2] Zhang H, Tian X, Lu X, et al. TMEM25 modulates neuronal excitability and NMDA receptor subunit NR2B degradation. J Clin Invest. 2019;129(9):3864-3876. doi:10.1172/JCI122599(IF:14.808)
[3] Wang T, Xiao G, Lu Q, et al. Synergistic Lysosomal Impairment and ER Stress Activation for Boosted Autophagy Dysfunction Based on Te Double-Headed Nano-Bullets. Small. 2022;18(27):e2201585. doi:10.1002/smll.202201585(IF:13.281)
[4] Wan SS, Zhang L, Zhang XZ. An ATP-Regulated Ion Transport Nanosystem for Homeostatic Perturbation Therapy and Sensitizing Photodynamic Therapy by Autophagy Inhibition of Tumors. ACS Cent Sci. 2019;5(2):327-340. doi:10.1021/acscentsci.8b00822(IF:12.837)
[5] Hu P, Li H, Sun W, et al. Cholesterol-associated lysosomal disorder triggers cell death of hematological malignancy: Dynamic analysis on cytotoxic effects of LW-218. Acta Pharm Sin B. 2021;11(10):3178-3192. doi:10.1016/j.apsb.2021.02.004(IF:11.614)
[6] Xiaoli F, Yaqing Z, Ruhui L, et al. Graphene oxide disrupted mitochondrial homeostasis through inducing intracellular redox deviation and autophagy-lysosomal network dysfunction in SH-SY5Y cells. J Hazard Mater. 2021;416:126158. doi:10.1016/j.jhazmat.2021.126158(IF:10.588)
[7] Wang N, Liu H, Liu G, et al. Yeast β-D-glucan exerts antitumour activity in liver cancer through impairing autophagy and lysosomal function, promoting reactive oxygen species production and apoptosis. Redox Biol. 2020;32:101495. doi:10.1016/j.redox.2020.101495(IF:9.986)
[8] Hu P, Wang J, Qing Y, et al. FV-429 induces autophagy blockage and lysosome-dependent cell death of T-cell malignancies via lysosomal dysregulation. Cell Death Dis. 2021;12(1):80. Published 2021 Jan 13. doi:10.1038/s41419-021-03394-4(IF:8.469)
[9] Ma WQ, Sun XJ, Zhu Y, Liu NF. PDK4 promotes vascular calcification by interfering with autophagic activity and metabolic reprogramming. Cell Death Dis. 2020;11(11):991. Published 2020 Nov 17. doi:10.1038/s41419-020-03162-w(IF:8.469)
[10] Peng J, Pan J, Wang H, Mo J, Lan L, Peng Y. Morphine-induced microglial immunosuppression via activation of insufficient mitophagy regulated by NLRX1. J Neuroinflammation. 2022;19(1):87. Published 2022 Apr 12. doi:10.1186/s12974-022-02453-7(IF:8.322)
[11] Liu Z, Lv X, Xu L, et al. Zinc oxide nanoparticles effectively regulate autophagic cell death by activating autophagosome formation and interfering with their maturation. Part Fibre Toxicol. 2020;17(1):46. Published 2020 Sep 18. doi:10.1186/s12989-020-00379-7(IF:7.546)
[12] Cui D, Ma J, Liang T, et al. Selenium nanoparticles fabricated in laminarin polysaccharides solutions exert their cytotoxicities in HepG2 cells by inhibiting autophagy and promoting apoptosis. Int J Biol Macromol. 2019;137:829-835. doi:10.1016/j.ijbiomac.2019.07.031(IF:6.953)
[13] Pan H, Wang Y, Na K, et al. Autophagic flux disruption contributes to Ganoderma lucidum polysaccharide-induced apoptosis in human colorectal cancer cells via MAPK/ERK activation. Cell Death Dis. 2019;10(6):456. Published 2019 Jun 11. doi:10.1038/s41419-019-1653-7(IF:5.959)
[14] Wang S, Xu X, Che D, et al. Reactive Oxygen Species Mediate 6c-Induced Mitochondrial and Lysosomal Dysfunction, Autophagic Cell Death, and DNA Damage in Hepatocellular Carcinoma. Int J Mol Sci. 2021;22(20):10987. Published 2021 Oct 12. doi:10.3390/ijms222010987(IF:5.924)
[15] Chen J, Wang D, Zong Y, Yang X. DHA Protects Hepatocytes from Oxidative Injury through GPR120/ERK-Mediated Mitophagy. Int J Mol Sci. 2021;22(11):5675. Published 2021 May 26. doi:10.3390/ijms22115675(IF:5.924)
[16] Kang L, Liu S, Li J, Tian Y, Xue Y, Liu X. The mitochondria-targeted anti-oxidant MitoQ protects against intervertebral disc degeneration by ameliorating mitochondrial dysfunction and redox imbalance. Cell Prolif. 2020;53(3):e12779. doi:10.1111/cpr.12779(IF:5.753)
[17] Wang D, Tan KS, Zeng W, et al. Hepatocellular BChE as a therapeutic target to ameliorate hypercholesterolemia through PRMT5 selective degradation to restore LDL receptor transcription. Life Sci. 2022;293:120336. doi:10.1016/j.lfs.2022.120336(IF:5.037)
[18] Kang L, Xiang Q, Zhan S, et al. Restoration of Autophagic Flux Rescues Oxidative Damage and Mitochondrial Dysfunction to Protect against Intervertebral Disc Degeneration. Oxid Med Cell Longev. 2019;2019:7810320. Published 2019 Dec 30. doi:10.1155/2019/7810320(IF:4.868)
[19] Xia Y, Xu C, Zhang X, et al. An activatable liposomal fluorescence probe based on fluorescence resonance energy transfer and aggregation induced emission effect for sensitive tumor imaging. Colloids Surf B Biointerfaces. 2020;188:110789. doi:10.1016/j.colsurfb.2020.110789(IF:4.389)
[20] Xia Z, Wang L, Li S, et al. ML-SA1, a selective TRPML agonist, inhibits DENV2 and ZIKV by promoting lysosomal acidification and protease activity. Antiviral Res. 2020;182:104922. doi:10.1016/j.antiviral.2020.104922(IF:4.101)
[21] Hou X, Han L, An B, et al. Mitochondria and Lysosomes Participate in Vip3Aa-Induced Spodoptera frugiperda Sf9 Cell Apoptosis. Toxins (Basel). 2020;12(2):116. Published 2020 Feb 13. doi:10.3390/toxins12020116(IF:3.531)
[22] Zhang X, Chen Q, Chen M, et al. Ambroxol enhances anti-cancer effect of microtubule-stabilizing drug to lung carcinoma through blocking autophagic flux in lysosome-dependent way. Am J Cancer Res. 2017;7(12):2406-2421. Published 2017 Dec 1. (IF:3.264)
[23] Gao Y, Xu A, Shen Q, Xie Y, Liu S, Wang X. Graphene oxide aggravated dextran sulfate sodium-induced colitis through intestinal epithelial cells autophagy dysfunction. J Toxicol Sci. 2021;46(1):43-55. doi:10.2131/jts.46.43(IF:2.196)